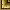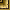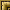




Л. С. Шалдыбин
Гельминтофауна млекопитающих мордовского государственного заповедника (Продолжение)
Ленточные черви
Семейство Anaplocephalidae Cholodkowsky, 1902.
Moniezia benedeni (Moniez, 1879).
Обнаружен единственный половозрелый экземпляр в кишечнике одного лося. Можно сказать, что лосиное поголовье по мониезиозу находится в относительно благополучном состоянии. Но вместе с этим нахождение М. benedeni у лося заповедника свидетельствует о том, что лоси могут быть хранителями, а следовательно, и источником этого заболевания. Это может быть, как и при дикроцелиозе, препятствием к полному уничтожению указанных инвазий в районе заповедника.
Mosgovoyta pectinata (Goeze, 1782).
Единственные экземпляры этого паразита обнаружены у трех зайцев-беляков. В нашей советской литературе (С. П. Наумов, 1947) отмечается, что интенсивность инвазии этим паразитом может быть очень высокая (до 83 экземпляров). Принимая во внимание литературные данные, можно сказать, что М. pectinata не является у зайца-беляка заповедника редкостью, но и не играет роли существенного отрицательного экологического фактора в жизни зайца-беляка.
Paranoplocephala отpholodes (Hermann, 1783).
Обнаружен в тонком кишечнике у двух взрослых водяных крыс в количестве 1—2 экземпляров.
Paranoplocephala brevis Kirschenblatt, 1938.
Обнаружен в тонком кишечнике одной обыкновенной полевки, одной полевки-экономки и у одной рыжей полевки.
Catenotaenia pusilla (Goeze, 1782).
Обнаружены единичные экземпляры в тонком кишечнике у 15 северных мышовок и у 9 рыжих полевок.
Семейство Hymenolepididae Furmann, 1907(Классификация семейства Hymenolepididae принимается по Скрябину и Матевосян, 1945).
Подсемейство Hymenolepidinae Perner, 1897.
Hymenolepis helenin. sp.
Обнаружен в тонком кишечнике трех кутор. Интенсивность инвазии от 1 до 3 экземпляров (рис. 4).

Рис. 4. Hymenolepis heleni n. sp.
Описание вида. Длина стробилы со сколексом 50 мм при максимальной ширине стробилы 1,2 мм. Длина гермафродитного членика 0,153 мм. Сколекс округлый, слегка вытянутый, с поперечником 0,350—0,510 мм. Хоботок вооружен одной короной из 46 крючьев, по форме близких к фратернойдному типу. Длина крючьев 0,033 мм. Присоски овальные, 0,272 мм в наибольшем поперечнике. Половые отверстия открываются посередине края членика. Семенники расположены в один ряд, плотно прилегая друг к другу, с диаметром 0,153 мм. Яичник лопастной, вместе с желточником занимает медианное положение. Бурса цирруса грушевидной формы, 0,159 мм длины при 0,085 мм максимальной ширины. Вооружение цирруса обнаружить не удалось. Наружный и внутренний семенные пузырьки хорошо развиты. Матка двухлопастная, мешковидная. Имевшиеся в нашем распоряжении экземпляры не имели зрелых члеников, поэтому размеры яиц остались нами не выясненными.
Дифференциальный диагноз
Описываемая форма по морфологии ближе всего стоит к Н. magnirostellata Baer, 1931 и Н. neomydis Baer, 1931, паразитирующим у куторы, но отличается от них размерами, количеством и формой крючьев, что дает нам право на описание нового вида.
Pseudobotrialepis niathewossiani Schaldybin, 1957 (рис. 5).

Рис. 5. Pseudobotrialepis malhewossiani n. g. n. s.
а - сколекс (общий вид), б - сколекс (апикально) в — гермафродитные членики.
Эти своеобразные цестоды были обнаружены в тонком кишечнике шести обыкновенных бурозубок и двух малых бурозубок. Интенсивность инвазии колебалась от 1 до 12 экземпляров. По обнаруженным экземплярам нами произведено первоописание вида.
Описание вида. Длина стробилы до 20 мм при максимальной ширине 1,1 мм. Членики на всем протяжении стробилы короткие, но сильно вытянутые в поперечном направлении. Длина гермафродитного членика 0,068—0,070 мм при ширине 0,6 мм. Шейка хорошо выражена. Сколекс сплюснут в дорзовентральном направлении, лишен хоботка и имеет 0,680 мм в поперечнике. По бокам сколекс имеет щелевидные углубления, напоминающие присасывательные ямы, на боковых поверхностях которых располагается по одной присоске, обращенной друг к другу и имеющей размер 0,255 x 0,204 мм. Семенники в количестве трех, неправильной овальной формы, с поперечником 0,050—0,068 мм расположены в одну линию. Два из них лежат апорально и один морально. Втянутый, слабо лопастной яичник и прилежащий к нему округлый желточник отделяют поральный семенник от двух апоральных. Бурса цирруса вытянутая, с хорошо развитыми внутренними семенными пузырьками, имеет 0,154 мм в длину при максимальной ширине 0,018—0,020 мм. Матка в зрелом состоянии мешковидная.
Dicranotaenia procera (Janicke, 1904).
Обнаружен в кишечнике одной водяной крысы в количестве трех экземпляров.
Dicranotaenia furcata (Stieda, 1862).
Обнаружен в кишечнике у 9 обыкновенных бурозубок и у четырех малых бурозубок в количестве от 1 до 22 экземпляров.
Vigisolepis spinulosa (Cholodkowsky, 1906).
Обнаружены единичные экземпляры у четырех обыкновенных бурозубок и у трех малых бурозубок.
В настоящее время существуют три различных описания этого вида. Одно описание сделано Холодковским в 1906 году, другое описание дает с в 1932 году и третье описание, под названием V. barbascolex, сделано Спасским в 1949 году. Анализируя дискуссию, изложенную Спасским при описании V. barbascolex, и изучив имевшиеся в нашем распоряжении материалы собственных сборов (18 экземпляров), мы пришли к выводу, что Холодковский и Спасский имели дело с одним и тем же видом — V. spinulosa.
Мы согласны с изменением диагноза рода Vigisolepis, которое привел Спасский, но не можем согласиться с описанием нового вида — V. barbascolex, которое он сделал, ссылаясь на различное число продольных полей крючьев и иную конфигурацию этих полей.
В отношении числа продольных полей крючьев Холодковский мог вполне ошибиться, так как на некоторых экземплярах действительно бывает трудно сделать заключение — 4 или 6 полей крючьев, потому что у таких экземпляров хоботок плохо просвечивает, и для подсчета полей необходимо прибегнуть к поворотам сколекса под микроскопом.
Что касается иной конфигурации продольных полей крючьев, то этот признак совершенно не может быть основанием к выделению нового вида, так как при просмотре серийного материала нетрудно убедиться в том, что эта конфигурация может варьировать. Поля крючьев могут быть короче и шире или длиннее и уже. Кольцевая зона мелких крючьев может быть составлена из 2, 3 и даже 4 рядов.
На основании вышеизложенного мы считаем, что описанием V. barbascolex Спасский дал более подробное и более точное описание V. spinulosa.
Что касается вида, который описал Baer под названием V. spinulosa, мы высказываем сомнение в его идентичности с V. spinulosa, описанным Холодковским.
Наше сомнение основано на там, что Baer видел всю поверхность хоботка покрытой крючьями, не выделяя продольных полей. Ошибиться в этом, изучая объект, имеющий продольные поля, нам кажется невероятным. Кроме этого, рисунок крюка из головной короны, по Baer (у), ясно отличается от такового, предложенного Холодковским.
Для окончательного решения этого вопроса необходим дополнительный материал. В противном случае этот вопрос решит время. Нахождение экземпляров, аналогичных описанию Baer (а), подтвердит самостоятельность вида;отсутствие повторных нахождений докажет справедливость диагноза Baer (а).
Подсемейство Pseudhymenolepidinae Joyeux et Baer, 1935.
До настоящего времени в литературном описании существует один род — Pseudhymenolepis Joyeus et Baer, 1935, с единственным видом — Pseudhymenolepis redonica, относящимся к этому подсемейству.
Причинами к выделению данного подсемейства явились:
1) Наличие у P. redonica матки, распадающейся на яйцевые капсулы, содержащие в себе по одному яйцу;
2) Наличие того факта, что членики отторгаются от стробилы на разной стадии своего развития и дальнейшее их развитие происходит изолированно, вне связи со стробилой.
В имеющемся у нас материале мы установили еще четыре вида, для которых также характерно распадение матки на яйценосные капсулы и отторжение недоразвитых члеников от стробилы. Нами установлено при этом, что отторжение члеников может происходить не только по одному, но и группами, образующими отдельные фрагменты, которые созревают вне связи со стробилой и в конечном своем развитии образуют единую капсулу с большим количеством яйцевых капсул, в нее включенных.
За последнее время в русской и иностранной литературе появилось описание еще ряда видов, имеющих аналогичные признаки.
Наличие двух основных признаков — матки, распадающейся на яйценосные капсулы, и отторжения недоразвитых члеников или групп члеников — относит наши формы к подсемейству Pseudhymenolepidinae, расширяя последнее.
В связи с последним мы считаем необходимым несколько расширить диагноз подсемейства.
Диагноз подсемейства Pseudhymenolepidinae no v. comb. Мелкие Hymenolepididae, у которых членики то одному или группами отторгаются от стробилы, будучи еще незрелыми. Дозревание этих члеников или фрагментов идет вне связи со стробилой и приводит к образованию общей капсулы, содержащей в себе яйцевые капсулы одного или несколько члеников, входящих в состав фрагмента.
Типичный род: Pseudhymenolepis Joyeux et Baer, 1936.
Neoskrjabinolepis stefanskii (Zarnowski, 1954)
Обнаружен в тонком кишечнике у трех обыкновенных бурозубок. Интенсивность инвазии от 3 до 15 экземпляров.
Вначале мы отнесли этот вид к роду Hymenolepis, но в дальнейшем в литературе появилось описание этого вида польским гельминтологом (Zarnowski) под названием Н. stefanskii.
В идентичности видов нет никакого сомнения, но польскому гельминтологу удалось подметить у этого вида образование из зрелого членика яйценосной капсулы, содержащей внутри себя множество капсул, каждая из которых содержит по 1 яйцу. Именно это обстоятельство и заставляет отнести этот вид к роду Neoskrjabinolepis, описанному Спасским в 1947 году.
Neoskrjabinolepis schaldybini Spassky, 1947
Обнаружен в тонком кишечнике у 22 обыкновенных бурозубок, у 8 малых бурозубок и у двух кутор (рис. 6). Интенсивность инвазии колебалась у обыкновенных бурозубок от 1 до 40 экземпляров, 7 малой бурозубки от 1 до 12 экземпляров и у куторы от 4 до 9 экземпляров.
Рис. 6. Neoskrjabinolepis schaldybini Spassky, 1947
а - сколекс, б - крюк, в - различные стадии созревания фрагмента
Изучение экземпляров этого вида привело нас к очень интересным наблюдениям. Мы подметили, что членики стробилы развиваются не каждый отдельно, а по нескольку одновременно. Благодаря этому в стробиле четко выделяются участки одинаковой зрелости. Далее, нами было отмечено, что фрагменты зреющих члеников отторгаются от стробилы и живут в кишечнике хозяина самостоятельно. При этом происходит распадение матки на яйценосные капсулы (в каждой капсуле по одному яйцу), после этого распадаются перегородки между члениками и образуется сложная яйценосная капсула. В конечном счете сложная капсула теряет кожно мышечную оболочку.
Образование капсулы с одним яйцом заставило нас отнести этот вид в подсемейство Pseudhymenolepidinae.
Своими соображениями мы поделились с А. А. Спасским. Спустя некоторое время Спасский опубликовал описание этого вида и назвал его нашим именем.
В своем описании Спасский трактует наблюдаемое нами явление иначе. Он считает, что имеется налицо не распадение матки на капсулы, а слияние маток члеников фрагмента.
Мы считаем это голословным и ошибочным утверждением. Доказательством нашей правоты служит наличие у яйца сложной капсулы не 3-х, а 4-х оболочек. Наружная оболочка и представляет собой производное стенки матки.
Учитывая все вышеизложенное, мы считаем необходимым изменить диагноз рода Neoskrjabinolepis, данный Спасским, и привести свое оригинальное описание вида.
Описание вида. Стробила очень нежная, почти прозрачная, 3—4 мм длиной при максимальной ширине 0,340 мм. Длина сколекса 0,272 мм при максимальной ширине 0,289—0,340 мм. Присоски округлые, 0,119—0,136 мм в диаметре. Хоботок вооружен десятью крючьями в одной короне. По форме крючья напоминают таковые Hymenolepis muttiatriata, но отличаются задним концом корня, который у данной формы в начальной своей части имеет пологий перехват, за которым следует широкоокруглый конец. Длина крючьев 0,039—0,052 мм. Стробила со сколексом не имеет при себе зрелых члеников. Всю стробилу можно разграничить по отделам. Первый отдел, самый узкий, следует непосредственно за сколексом, не имеющий поперечной исчерченности, — шейка. За шейкой следует небольшой поперечноисчерченный отдел — отдел образования проглотид. Третий отдел из 5—25 проглотид, каждая из которых имеет формирующийся или уже сформировавшийся гермафродитный половой аппарат одинаковой степени зрелости. Четвертый отдел состоит из 5—25 зреющих проглотид. Как и в предыдущем отделе, степень зрелости одинаковая во всех проглотидах данного отдела. Переходы от отдела к отделу не постепенные, а резкие. Промежуточных стадий зрелости между отделами нет. Гермафродитный членик всегда очень короткий, но широкий. Длина такого членика 0,015 мм при ширине 0,170—0,210 мм. Семенники расположены в один ряд, по типу VII — женские железы отделяют поральный семенник от двух апоральных. Семенники неправильно округлой формы, с диаметром 0,012 мм. Желточник слаболопастной. Бурса цирруса вытянутая, волнообразно изогнутая, 0,050—0,055 мм длины. Циррус вооружен шипиками. Матка мешковидная, распадающаяся при созревании яиц на капсулы, содержащие по одному яйцу. Четвертый отдел остается при общей части стробилы до момента распада матки на капсулы. После отторжения конечного отдела (фрагмента) стробилы последний ведет самостоятельный образ жизни в кишечнике хозяина.
Дальнейшее развитие фрагмента состоит в том, что яйца дозревают, увеличиваются в размере. При этом и весь фрагмент увеличивается в размере, достигая до 0,510 мм в ширину. Одновременно уменьшается паренхиматозный слой и происходит резорбция перегородок между проглотидами, благодаря чему яйцевые капсулы из всех проглотид оказываются вместе, включенные в бесструктурную массу. Наружные границы отдельных проглотид остаются четко заметными, как на стробиле. Вслед за этим кожно-мышечная стенка фрагмента трескается по границам проглотид и по частям спадает с фрагмента. После исчезновения кожно-мышечного слоя («чехлика») остается фрагмент, по форме напоминающий первоначальный, но значительно тоньше и состоящий из совершенно прозрачной, гомогенной массы, в центре которой сконцентрированы яйцевые капсулы в количестве нескольких сот. Подробное изучение этих конечных образований показало, что они не имеют в себе ни полости, ни каналов, ни отверстий. На концах, по которым яйцевые капсулы могли бы покидать их. При легком надавливании на фрагмент, яйцевые капсулы с одинаковой легкостью во всех направлениях могут проходить свободный гомогенный слой и выходить через него в любом месте фрагмента. Это доказывает полное отсутствие специальных выводных протоков. Яйцевая капсула совершенно округлая, 0,051 мм в диаметре. За оболочкой капсулы следует очень нежная, часто складчатая, оболочка яйца. Онко-сфера имеет 0,021 мм в диаметре.
Диагноз рода Neoskrjabinolepis Spassky 1947 n. comb. Pseud-hymenolepidinae, сколекс которых вооружен одной короной крючьев. Половые отверстия односторонние. Три семенника расположены в одну поперечную линию. Женские железы лежат медианно. Матка мешковидная, распадающаяся на яйцевые капсулы, содержащие в себе по одному яйцу. Задний созревающий членик или ряд созревающих члеников отторгаются и развиваются дальше вне связи со стробилой. В отторгнувшемся фрагменте члеников межпроглотидные перегородки резорбируются, а яйцевые капсулы оказываются лежащими в гомогенной прозрачной массе, сохраняющей форму фрагмента после слущивания кожно-мышечного слоя. Паразиты насекомоядных. Типичный вид: N. schaldybini Spassky, 1947.
Spasskylepis ovaluteri n, gen. n. sp. (рис. 7).
Обнаружен в тонком кишечнике у семи обыкновенных бурозубок. Интенсивность инвазии колебалась от 1 до 10 экземпляров.

Рис. 7. Spasskylepis ovaluteri n. gen. n. sp.
Описание вида. Мелкие цестоды, 4—7 мм длины и 0,221—0,316 мм ширины. Сколекс 0,153 мм длины при максимальной ширине 0,170 мм, лишен хоботка, сплюснут в дорзовентральном направлении. Овальные присоски 0,153 x 0,084 мм размером расположены по две с дорзальной и вентральной сторон, плотно прилегая друг к другу. С боковой стороны сколекс напоминает шляпку гриба. Стробила имеет четыре отдела: шейку, отдел формирования члеников, отдел гермафродитных члеников и отдел зреющих члеников. Гермафродитные членики 0,068 мм длины и 0,187 мм ширины. Проглотиды зреющего отдела имеют длину, близкую к ширине или превышающую ее. Три округлых семенника 0,051 мм в диаметре расположены треугольником. Передний семенник лежит медианно перед двумя задними. Яичник и желточник занимают центральное положение. Бурса цирруса вытянутая, 0,114 мм длины. Циррус не вооружен. Матка мешковидная, круглая или овальная, распадающаяся на капсулы, содержащие по одному яйцу. Зрелые членики в количестве 5—25 отторгаются от стробилы и ведут самостоятельный образ жизни в кишечнике хозяина, формируя фрагмент, состоящий из гомогенной прозрачной массы с включенными в нее яйцевыми капсулами. По морфологии этот фрагмент не отличается от такового других видов данного подсемейства.
Диагноз рода Spasskylepis n. gen. Мелкие цестоды, у которых в каждом членике по три семенника, расположенных треугольником. Сколекс невооруженный. Членики по одному или группами отторгаются от стробилы и развиваются в дальнейшем вне связи со стробилой. У видов с групповым отторжением члеников три дальнейшем развитии происходит распадение матки на яйцевые капсулы, содержащие по одному яйцу. Паразиты насекомоядных.
Ditestolepis diaphana (Cholodkowsky, 1906). (рис. 8—10)

Рис. 8. Ditestolepis diaphana (Cholodkowsky, 1906)
(а — сколекс, б — общий вид)

Рис. 9. Ditestolepis diaphana (Cholodkowsity, 1906)
в - членики различной зрелости

Рис. 10. Ditestolepis diaphana (Cholodkowsky, 1906)
г — сложная капсула, д — одно яйцо в капсуле
Обнаружен в тонком кишечнике у 68 обыкновенных бурозубок, 17 малых бурозубок и у одной куторы. Интенсивность инвазии у бурозубок колебалась от 1 до 260 экземпляров.
Описание вида(по собственному материалу).
Мелкие цестоды, 2—4 мм длины и 0,280—0,350 мм максимальной ширины. Сколекс 0,160 мм длины при ширине 0,170 мм, лишен хоботка и сдавлен в дорзовентральном направлении. Овальные присоски 0,084 x 0,153 мм размером расположены по две с дорзальной и вентральной сторон близко друг к другу. С латеральной стороны сколекс напоминает шляпку гриба. Стробила имеет 4 отдела: шейку, отдел формирования проглотид, отдел гермафродитных члеников и отдел зреющих члеников. Переход от отдела к отделу сопровождается резким увеличением ширины и длины члеников. Третий и четвертый отделы включают в себя от 5 до 25 члеников, чаще 16—20. Все членики отдела одинаковы по степени зрелости. Семенники в количестве двух расположены по поперечной линии, округлые и имеют 0,036 мм в диаметре. Яичник слаболопастной. Бурса вытянутая, 0,105 мм длины, с хорошо развитым внутренним семенным пузырьком. Циррус не вооружен. Матка мешковидная, двухлопастная, распадающаяся на яйцевые капсулы, содержащие в себе по одному яйцу. Гермафродитный членик 0,042—0,051 мм длины и 0,153 мм ширины. Зреющий отдел отторгается от стробилы и превращается в единый фрагмент из прозрачной гомогенной массы, содержащей в себе несколько сотен яйцевых капсул.
Чрезвычайно мелкие, прозрачные цестоды, 0,393—0,459 мм длины вместе со сколексом. Сколекс, лишенный хоботка, сплюснут в дорзовентральном направлении и несет на себе овальные присоски, расположенные по две с дорзальной и вентральной сторон, 0,084 x 0,066 мм размером. Вся стробила состоит из хорошо выраженной шейки и 5—6 члеников. Гермафродитный членик 0,039 мм длины при ширине 0,130 мм. Семенники в количестве 2 лежат поперек членика. Вентральнее семенников медианно располагается слаболопастной яичник. Половая бурса 0,054 мм длины с хорошо развитым внутренним семенным пузырьком. Циррус не вооружен. Матка лопастная, мешковидная, распадающаяся на яйцевые капсулы, содержащие по одному яйцу. Зреющий членик отторгается от стробилы и созревает самостоятельно, образуя фрагмент, лишенный кожно-мышечного слоя и содержащий в себе яйцевые капсулы диаметром 0,051 мм.
Дифференциальный диагноз
По своему строению описываемая цестода очень близка с D. diaphana, но отличается значительно меньшим размером и тем, что стробила лишена резко выраженных отделов. Стробила описываемой формы состоит всего из 5—6 члеников, последовательно развитых. Стробила же D. diaphana состоит из большого количества члеников и подразделяется но степени развитости члеников на 4 отдела. В отличие от D. diaphana описываемая форма не имеет явления слияния зрелых члеников.
Впервые род Ditestolepis был выделен Солтисом в 1952 году. Солтис, как и Спасский, считает, что при образовании «синкапсулы» происходит слияние маток, а не распадение их на отдельные капсулы. Мы с этим согласиться не можем и вносим изменение в диагноз рода.

Рис. 11. Ditestoleois secunda n. sp.
Диагноз рода Ditestolepis Soltys, 1952 п. comb. Цестоды (Нуmenolepididae) небольших размеров, с невооруженным сколексом. Членики, вытянутые в поперечном направлении, закладываются в области шейки поодиночке или сериями. В последнем случае стробила имеет ступенчатое строение. Половой аппарат непарный, половые поры односторонние. В каждом членике образуется по два семенника, расположенных по сторонам от женских половых желез, залегающих по средней линии. Матка вначале мешковидная. При созревании яиц матка распадается на отдельные капсулы, содержащие в себе по одному яйцу. При серийном созревании члеников последние сливаются вместе, образуя единый фрагмент, лишенный кожно-мышечного слоя. Паразиты насекомоядных.
Помимо указанных в нашей работе родов, к подсемейству Pseudhymenolepidinae следует отнести и такие известные в литературе роды, как Protogynella Iones, 1943 и Soricinia Spassky, et Spasskaja, 1954.
Семейство Dilepididae Furmann, 1907.
Choanotaenja crassiscolex (Lipstow, 1890).
Обнаружен в тонком кишечнике 75 обыкновенных бурозубок, б малых бурозубок и у 5 кутор. Интенсивность инвазии достигала до 114 экземпляров.
Семейство Taeniidae Ludw., 1886.
Taenia hydatigena (Pall., 1766).
Обнаружен в тонком кишечнике 14 волков. Интенсивность инвазии колебалась от 1 до 13 экземпляров.
Массовая инвазия волка этой цестодой делает его основным источником цистицеркоза диких жвачных животных заповедника.
Taenia polyacantha Leuckart, 1856.
Обнаружен в тонком кишечнике 2 волков по 1 экземпляру.
Taenia crabbei Moniez, 1879.
Обнаружен в тонком кишечнике 1 волка. Интенсивность инвазии 1 экземпляр.
Эта цестода является типичным паразитом песцов, волков, собак и кошек северной части СССР, где в качестве промежуточного хозяина выступает северный олень. В Мордовском заповеднике Т. crabbei регистрируется нами не первыми. В 1946 году этот паразит был обнаружен в числе нескольких экземпляров Ф. Н. Морозовым в материалах, полученных от заповедника из волков.
В стадии цистицерка Т. crabbei нами в заповеднике не зарегистрирован. Вполне возможно, что заражение волков этим паразитом происходит в более северных районах. Это свидетельствует о широкой способности волка к миграциям, а следовательно, и о большой роли его в распространении инвазии.
Multlceps multlceps (Leske, 1780).
Обнаружен в тонком кишечнике у 6 волков и 1 лисицы. Интенсивность инвазии у волка колебалась от 1 до 10 экземпляров, а у лисицы 3 экземпляра.
Multiceps serialis (Garwais, 1847).
Обнаружен в тонком кишечнике у 4 волков и двух лисиц. Интенсивность инвазии у волков колебалась от 1 до 65 экземпляров, у лисиц — от 1 до 77 экземпляров.
Семейство Mesocestoididae Furmann, 1907.
Mesocestoideslineatus (Goeze, 1782).
Обнаружен в тонком кишечнике трех волков. Интенсивность инвазии 1—2 экземпляра.
Слабая инвазия волка этой цестодой объясняется тем, что волк редко поедает мышевидных грызунов, а последние являются, промежуточными хозяевами в развитии М. lineatus.
Круглые черви
Семейство Trichostrongylidae Leiper, 1912.
Trichostrongylus retortaeformis (Zeder, 1800).
Обнаружен в тонком кишечнике 14 зайцев-беляков. Интенсивность инвазии колебалась от 1 до 2844 экземпляров, в среднем 754 экземпляра на 1 зайца.
Trichostrongylus colubriformis (Giles, 1892).
Обнаружен в тонком кишечнике 3 зайцев-русаков. Интенсивность инвазии колебалась от 12 до 529 экземпляров.
Ostertagia ostertagl (Stiles, 1892).
Обнаружен в сычуге и тонком кишечнике у 11 лосей и 1 марала. Наибольшая интенсивность инвазии наблюдалась у лося, где она колебалась от 1500 до 15568 экземпляров.
Spiculopteragia alcis. Schulz, Kadenazii, Evranova et Schaldybin, 1954 (рис. 12 и 13).
Обнаружен в сычуге и тонком кишечнике всех исследованных лосей. Интенсивность инвазии, как и предыдущим видом, высокая, колебалась от 700 до 7784 экземпляров.

Рис. 12. Spiculopteragia alcis. Schulz, Kadenazii, Evranova et Schaldybin, 1954 (Бурса)

Рис. 13. Spiculopteragia alcis. Schulz, Kadenazii, Evranova et Schaldybin, 1954 (Спикула)
S. alcis как новый вид стал известен четырем исследователям: Шульцу, Каденации, Эврановой и Шалдыбину. Эти товарищи одновременно и независимо друг от друга в разных зонах СССР обнаружили эту нематоду у лося.
Описание S. alcis было впервые опубликовано в 1954 по материалу Шульца и Каденации.
В настоящей работе мы приводим описание по своему оригинальному материалу.
Описание вида. Самец 7,65—8,30 мм длины и 0,119—0,136 мм максимальной ширины (перед бурсой). Ширина на уровне конца пищевода 0,035—0,102 мм. Диаметр головного конца 0,021— 0,024 мм. Пищевод 0,680 мм длины и 0,042—0,054 мм ширины (в конечной своей части). Нервное кольцо окружает пищевод на расстоянии 0,270—0,303 мм от головного конца. Шейные сосочки располагаются на расстоянии 0,430 мм от головного конца. Бурса состоит из двух мощных латеральных и одной маленькой дорзальной лопастей. Ребра бурсы такие же, как и у других представителей рода. Некоторое отличие составляет дорзальное ребро, которое, хотя и массивное, но относительно длинное. На своем суживающемся конце дорзальное ребро бифурцирует, и каждая веточка в свою очередь разделяется на две коротких веточки.
Спикулы 0,172—0,187 мм длины, массивные и компактные. В дистальной трети каждая спикула расщеплена на два отростка. Внутренний отросток несколько короче наружного и заканчивается шаровидным утолщением. Наружный отросток значительно больше, серпообразно изогнут и на своем конце снабжен небольшим веерообразным расширением. Губернакулум отсутствует.
Самка 6,936—9,265 мм длины и 0,102—0,117 мм максимальной ширины (в области вульвы). Диаметр головного конца 0,021—0,030 мм. Ширина в области конца пищевода 0,102 мм. Пищевод 0,700—0,731 мм длиной при ширине 0,039—0,042 мм. Шейные сосочки расположены от головного конца на 0,342 мм. Вульва открывается поперечной щелью на расстоянии 1,479— 1,955 мм от хвостового конца и кутикулярного клапана не имеет. Хвостовой конец заканчивается небольшим закруглением. Анус открывается на расстоянии 0,221 мм от хвостового конца. Яйцемет имеет вид вытянутого боченка. Яйца 0,060—0,063 мм длины и 0,027—0,033 мм ширины.
Spiculopteragia panticola nov. sp.
Обнаружен в сычуге у пяти пятнистых оленей. Интенсивность инвазии не превышала 20 экземпляров (рис. 14 и 15).

Рис. 14. Spiculopteragia panticola n. sp. (Бурса)

Рис. 15. Spiculopteragia panticola n. sp.(Спикулы).
Описание вида. Самец 5,525—6,290 мм длины и 0,081—0,120 мм максимальной ширины (перед бурсой). Диаметр головного конца 0,066 мм. Пищевод 0,570—0,600 мм длины и 0,066 мм ширины в области своего конца. Шейные сосочки на расстоянии 0,286—0,300 мм от головного конца. Пребурсальные сосочки есть. Бурса напоминает таковую S. schulzi Rajewsk. 1930 и состоит из двух латеральных лопастей и маленькой дорзальной, которая с боков частично поддерживается массивными наружно-дорзальными ребрами, а посередине—дорзальным ребром. Дорзальное ребро относительно мощное, образует бифуркацию. Каждая ветвь в свою очередь в последней своей трети раздваивается. Вентро-вентральное ребро тоньше латеровентрального. Из латеральных ребер наружное латеральное переднее латеральное одинаковы и сближены друг с другом;задне-латеральное ребро отделено от остальных, тоньше и имеет по своему-ходу расширение. Спикулы одинаковые, светло-желтого цвета: 0,165—0,174 мм длины. На своем дистальном конце спикулы расщеплены на два отростка. Оба отростка заострены и шилообразно изогнуты во внутреннюю сторону. Между отростками располагается тонкая, слегка желтоватая перепонка. Вентральный отросток снабжен продолговатым веерообразным расширением. Губернакулум отсутствует. Самка 8,160—9,306 мм длины при максимальной ширине 0,105—0,170 мм (в области вульвы). Диаметр головного конца 0,027 мм. Пищевод 0,561—0,582 мм длины и 0,045 мм ширины в конечной своей части. Экскреторная пора располагается на 0,240—0,246 мм от головного конца. Вульва в виде поперечной щели располагается на 1,360 мм от головного конца. Зрелых яиц обнаружить не удалось.
Дифференциальный диагноз
От всех видов рода описываемая форма отличается главным образом наиболее простым устройством спикул. Спикулы у всех видов этого рода в своей диетальной части имеют два отростка, из которых медиальный всегда короче латерального, и по форме оба отростка разные.
У описываемой формы отростки одинаковой конфигурации, шиловидно загнуты внутрь и соединены тонкой перепонкой. Разница между отростками состоит только в том, что наружный из них имеет веерообразное расширение.
Molinostrongylus skrjabini Skarbilovitch, 1934.
Обнаружен в кишечнике у десяти рыжих вечерниц и у одной прудовой ночницы. Интенсивность инвазии у рыжей вечерницы достигала 129 экземпляров, а у прудовой ночницы равнялась 12 экземплярам.
Nematodirella longispiculata Jorke et Maplestone, 1926.
Обнаружен в тонком кишечнике у девяти лосей и у одного пятнистого оленя. Интенсивность инвазии у лосей достигала 4604 экземпляров, а у пятнистого оленя — 1 экземпляр.
Семейство StrongyHdae Baird, 1853.
Oesophagostomum asperum Railliet et Henry, 1913.
Обнаружен в толстом кишечнике у двух лосей и у одного пятнистого оленя. Интенсивность инвазии у лося не превышала 15 экземпляров, у оленя один экземпляр.
Oesophagostomum radlatum (Rud., 1863).
Обнаружен в толстом кишечнике четырех пятнистых оленей. Интенсивность инвазии колебалась от 1 до 10 экземпляров.
Семейство Heligmosomatidae Cremt 1927.
Heligmosomum skrjabini Schulz, 1926.
Обнаружен в тонком кишечнике у трех желтогорлых мышей и у одной лесной мыши. Интенсивность инвазии у желтогорлой мыши колебалась от 1 до 609 экземпляров, а у лесной мыши 25 экземпляров.
Heligmosomum polygirum (Duj, 1845).
Обнаружен в тонком кишечнике у десяти рыжих полевок и у двух пашенных полевок. Интенсивность инвазии не превышала 12 экземпляров.
Longi striata wolgensis Schulz, 1926.
Обнаружен в тонком кишечнике у четырех рыжих полевок и у двух пашенных полевок. Интенсивность инвазии достигала 46 экземпляров.
Longistriata paradox nov. sp.
Обнаружен в тонком кишечнике у 67 обыкновенных бурозубок, у 16 малых бурозубок и у 3 кутор. Наибольшая интенсивность инвазии достигала у обыкновенной бурозубки — 213 экземпляров (рис. 16 и 17).

Рис. 16. Longistriata paradox nov. sp.
(головной и хвостовой концы самки)

Рис. 17. Longistriata paradoxi nov. sp. (Бурса)
Описание вида. Самец 1,360—1,394 мм длины при максимальной ширине 0,042—0,051 мм. Диаметр головного конца 0,012 мм. Ширина в области конца пищевода 0,033—0,039 мм. Ширина перед бурсой 0,027 мм. Головной конец имеет кутикулярное вздутие 0,045—0,048 мм длиной и 0,030 мм шириной. По телу тянется шесть продольных гребней. Расстояние между продольными гребнями 0,018 мм. Цервикальные сосочки отстоят от головного конца на 0,045—0,048 мм. Пищевод 0,120—0,135 мм длины при ширине 0,030 мм в своем дистальном конце. На границе второй и последней трети своей длины пищевод окружен нервным кольцом. Половая бурса 0,075—0,099 мм длины при ширине 0,155—0,165 мм (в развернутом состоянии). Дорзальное ребро мощное и длинное, в конечной своей части разделяется на две короткие ветви, которые и свою очередь делятся вновь на две ветви. Своими концами ветви дорзального ребра доходят до края бурсы. Наружно-дорзальные ребра, мощные в своем основании и резко суживающиеся к концу, отходят асимметрично от дорзального ребра, (правое наружно-дорзальное ребро отходит от дорзального ребра несколько позже, чем левое). Латеральные ребра имеют мощный общий ствол, разделяющийся на три соответствующих ребра лишь в начале второй половины общей длины. Все три латеральных ребра имеют коническую форму с заостренными концами.
Самка 1,700—1,955 мм длины при максимальной ширине во второй половине тела 0,063—0,085 мм. Диаметр головного конца 0,012 мм, ширина в области конца пищевода 0,030—0,048 мм в области вульвы 0,048—0,075 мм. Головное кутикулярное вздутие 0,039—0,048 мм длины при ширине 0,033—0,036 мм. Хвостовой конец тупо закруглен. Анальное отверстие располагается субтерминально. Вульва открывается с дорзальной стороны и располагается на 0,069— 0,090 мм от хвостового конца. Яйца, 0,060 x 0,030 мм размером, располагаются одно за другим в один ряд.
Дифференциальный диагноз
Описываемый вид отличается от всех многочисленных видов данного рода своим своеобразным строением половой бурсы самца. Своеобразие заключается в том, что латеральные ребра начинаются массивным общим стволом, расщепляющимся на отдельные ребра лишь в начале второй половины общей длины, а вентральные ребра отходят от ствола латеральных ребер.
Этот признак резко отличает описываемую форму от всех других этого рода.